提到东北大米,除了软糯香甜的口感,其强悍的耐寒能力更令人称道——能在东北寒冷的春季扎根生长,抵御低温胁迫,而同为水稻的籼稻却难以适应低温环境。这种差异背后,藏着一套精密的分子调控体系。
水稻耐寒是多基因、多通路协同调控的复杂性状。种康院士团队深耕多年,一系列关键文献逐步勾勒出粳稻应对低温的“生存策略”——从上游的低温信号感知,到中游的代谢网络调控,再到下游的损伤修复,为水稻耐寒育种提供了坚实的分子基础。
一、COLD1基因,粳稻耐寒的“信号开关”
低温胁迫来临时,植物首先要完成“低温识别”——这是耐寒应答的第一步,也是最关键的起点。2015年,种康院士团队鉴定了负责低温感知的核心基因COLD1 [1],破解了粳稻与籼稻耐寒差异的源头密码,这一发现成为解析水稻耐寒机制的重要突破。 该研究通过正向遗传学筛选,从粳稻背景中鉴定出低温敏感突变体cold1-1,该突变体因COLD1 基因缺失全长转录本,无法正常感知低温。进一步实验证实,COLD1蛋白主要定位于内质网和细胞膜,这两个位置是植物感知外界环境信号的“前沿阵地”。
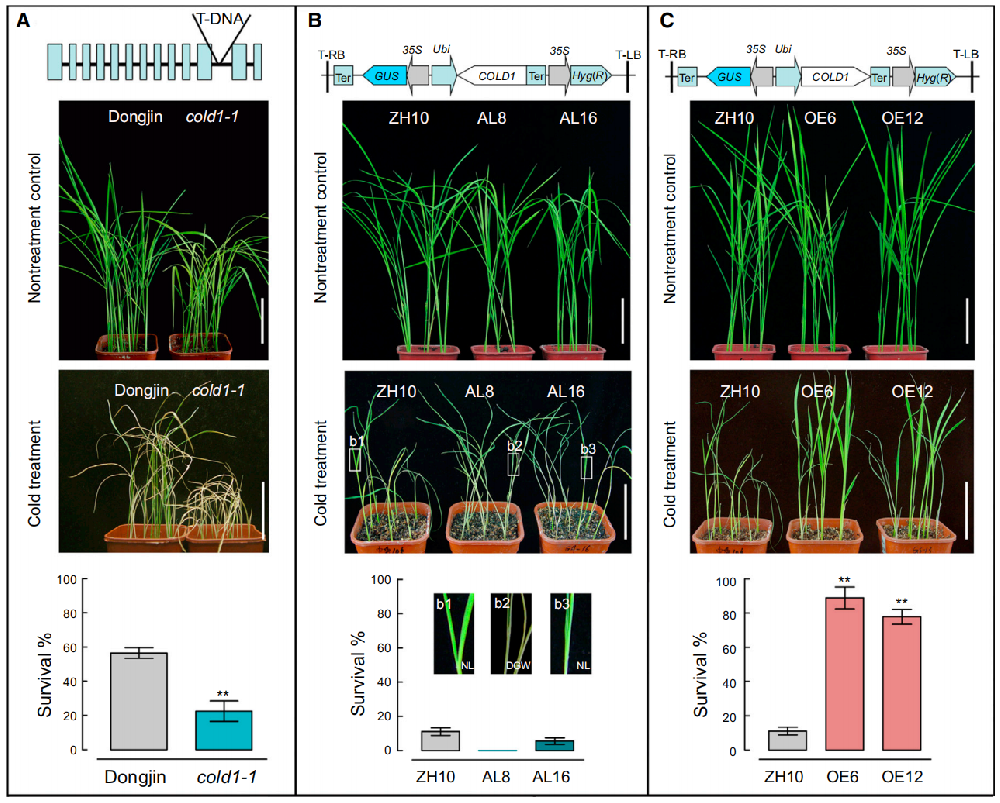
图1 COLD1调控水稻耐寒性[1]
(A:cold1-1 突变体表现出耐寒敏感性。B:反义转基因水稻株系(AL8 和 AL16)表现出耐寒敏感性。C:过表达转基因水稻株系(OE6 和 OE12)表现出耐寒性。)
研究发现粳稻与籼稻的COLD1 基因存在一个关键差异位点——SNP2。粳稻的SNP2位点为A碱基,编码赖氨酸(Lys¹⁸⁷);而籼稻的SNP2位点为T/C碱基,编码甲硫氨酸(Met¹⁸⁷)或苏氨酸(Thr¹⁸⁷)。这个单一氨基酸的替换,直接决定了COLD1蛋白的功能差异。
遗传互补实验显示,将粳稻型COLD1(SNP2ᴬ)转入cold1-1 突变体,能完全恢复其耐寒性;而转入籼稻型COLD1(SNP2ᵀ/ᶜ)则无法恢复。进化分析进一步表明,粳稻的COLD1 耐寒等位基因起源于中国普通野生稻,在粳稻驯化过程中被人工选择保留,成为粳稻适应北方低温环境的核心“遗传印记”。
COLD1的感知机制并不孤立:它能与水稻G蛋白α亚基RGA1物理相互作用,作为GTP酶激活因子加速RGA1的GTP水解活性,进而触发下游的钙离子(Ca²⁺)内流——这是低温信号从“感知”到“传导”的关键一步,为后续的耐寒应答打开了信号通道。
二、维生素E-维生素K1调控,耐寒差异的“核心枢纽”
当COLD1 感知低温并传递信号后,粳稻需要启动下游的代谢调控来抵御低温损伤。该团队在2021年的研究[2]聚焦COLD1 下游的代谢网络,发现叶绿体中维生素E-维生素K1子网络是水稻耐寒性差异的关键,完善了低温信号传导后的防御通路。
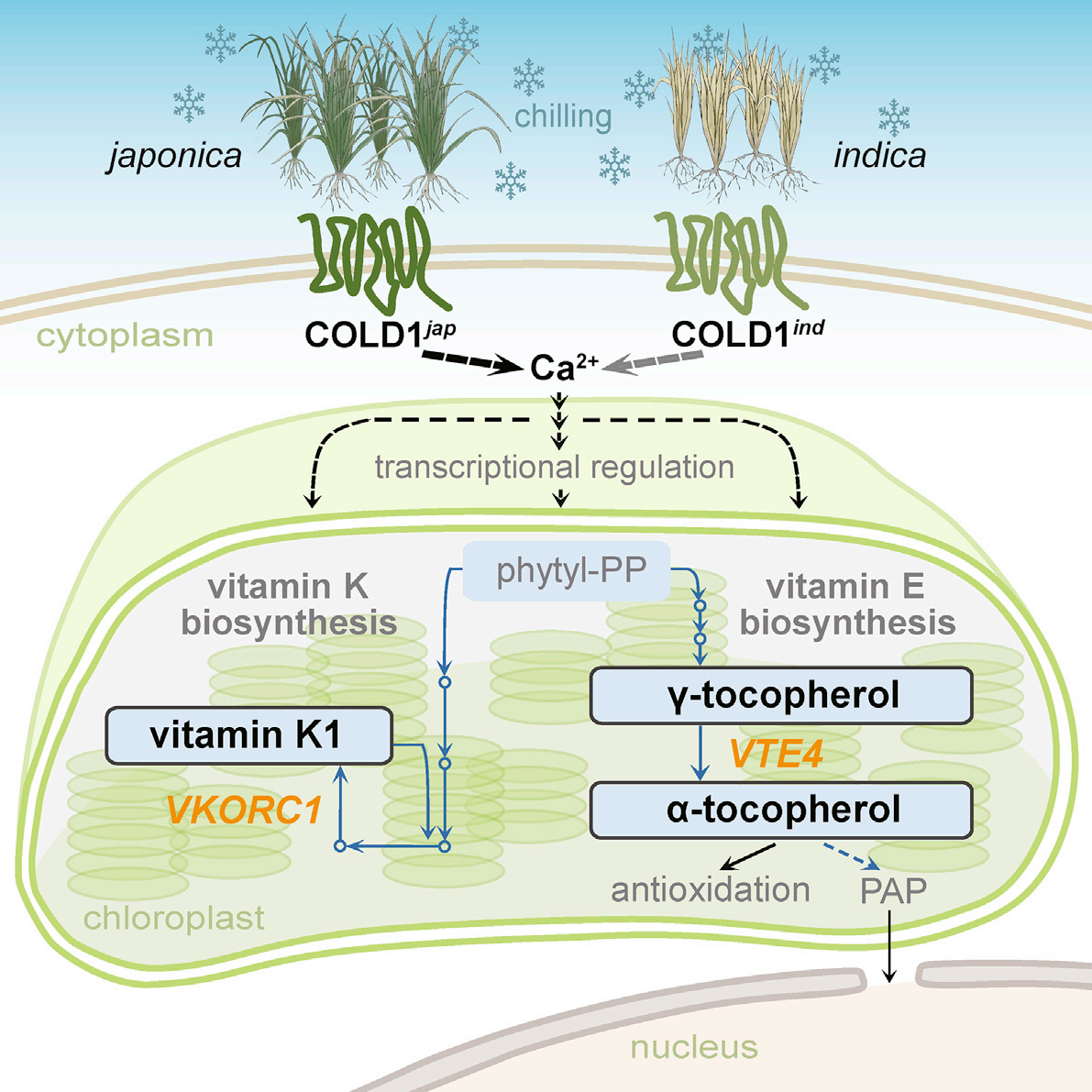
图2 维生素E -维生素K1子网络调控粳稻耐寒的模式图[2]
叶绿体是植物光合作用的场所,也是低温胁迫的敏感部位——低温会破坏叶绿体膜结构,抑制光合作用,还会诱发活性氧积累,造成细胞损伤。而维生素E(生育酚)和维生素K1(叶绿醌),正是叶绿体膜的“保护者”和活性氧的“清除者”。 研究通过染色体片段置换系(CSSL)和转基因技术验证:粳稻中COLD1 的激活会上调维生素E合成关键基因VTE4 的表达,显著增加α-生育酚(维生素E活性最强的亚型)含量;同时促进维生素K1的合成积累。而籼稻中,因COLD1功能较弱,VTE4 表达量低,维生素E/K1含量不足,叶绿体膜更易受低温破坏。 进一步实验发现,过表达VTE4 的籼稻植株,α-生育酚含量提升,耐寒性显著增强;而VTE4突变体(vte4)则表现为低温敏感,叶绿体膜损伤严重、叶绿素降解加快。这说明,COLD1通过调控维生素E/K1代谢,为粳稻构建了一道“叶绿体保护屏障”。 此外,研究还通过转录组和代谢组联合分析,勾勒出COLD1 调控的下游代谢网络:除了维生素代谢,COLD1还会协调碳水化合物代谢、脂质代谢,共同维持低温下粳稻的细胞稳态和生理功能——比如促进可溶性糖积累,提升细胞渗透压,抵御低温导致的细胞脱水。
三、COLD11介导的DNA修复,完善耐寒全链条
即便有了信号感知和代谢防御,低温仍可能会造成DNA损伤(如双链断裂DSB),若无法及时修复,会导致基因突变、细胞死亡,最终影响水稻存活。该团队2023年的研究[3]发现了参与寒害DNA损伤修复的基因模块,其中COLD11是核心成员,为粳稻耐寒机制补上了“损伤修复”的关键一环,与此前的信号感知、代谢调控研究形成互补。 该研究通过创新的数据融合全基因组关联分析(GWAS),突破了粳稻与籼稻表型极化的分析难题,成功定位到新的耐寒QTL位点qCTS11-1,其候选基因为COLD11 ——该基因编码RAD51A1蛋白,是DNA双链断裂修复的关键因子。
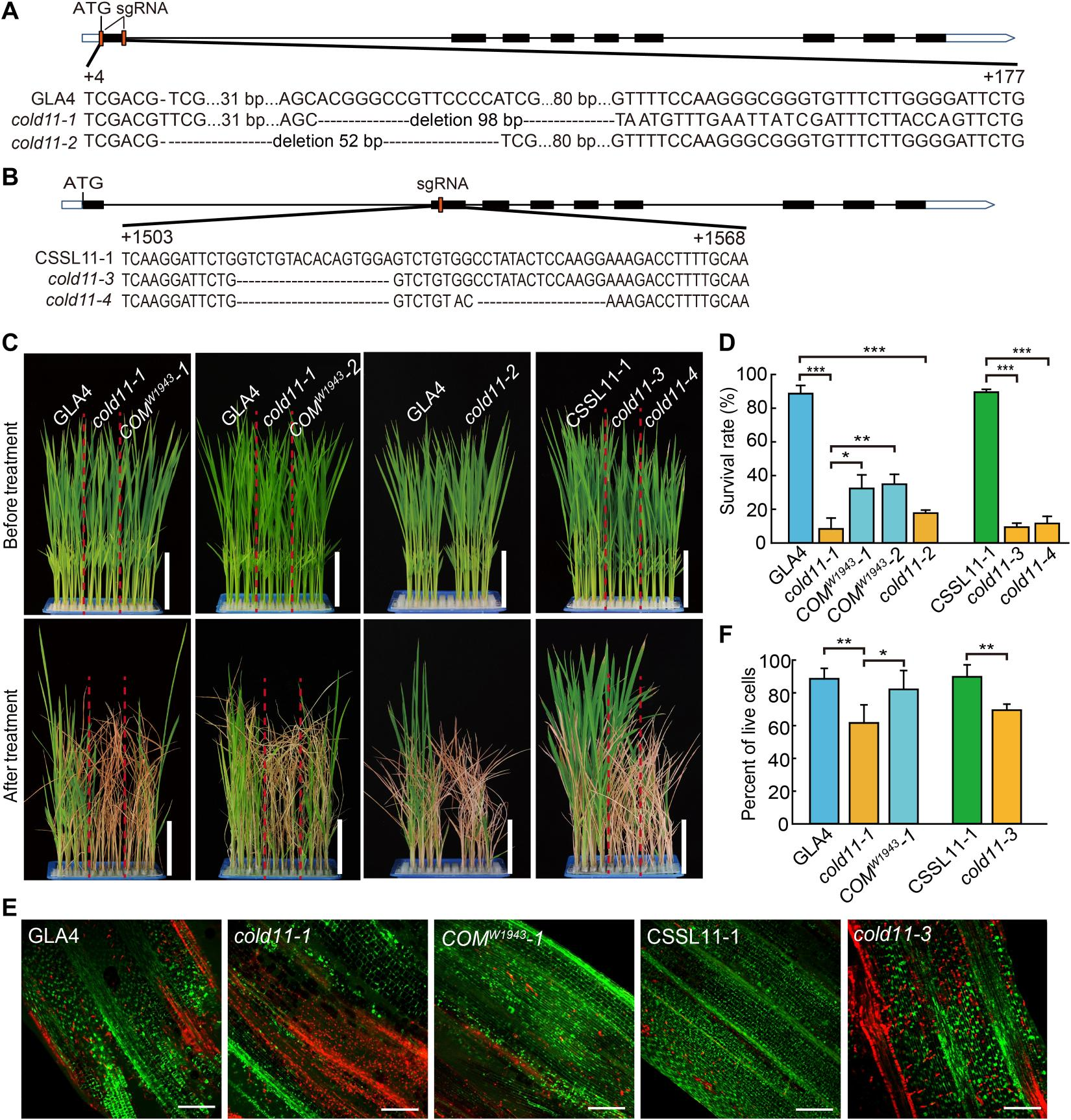
图3 COLD11 耐寒性验证[3]
实验证实,COLD11基因的功能缺失突变体(cold11-1、cold11-2等),低温处理后DNA双链断裂数量显著增加,细胞死亡率升高,耐寒性大幅下降;而互补实验能恢复其修复能力和耐寒性。这表明,COLD11是粳稻应对低温DNA损伤的“修复工具”。
COLD11的功能差异同样源于基因序列的自然变异——其第一外显子中GCG密码子(编码丙氨酸)的重复次数,直接影响修复能力。粳稻(尤其是温带粳稻,如东北大米)的COLD11含有更多GCG重复(通常超过3个),而籼稻多为3个重复。
研究发现,GCG重复次数越多,COLD11蛋白形成二聚体的能力越强(二聚体是RAD51家族蛋白发挥DNA修复功能的必需形式),DNA修复效率越高。进化分析显示,GCG重复次数的增加的是粳稻驯化过程中被选择的结果,与粳稻向北方低温地区扩张密切相关。
更重要的是,COLD11与COLD1并非独立作用——低温下,COLD1感知信号并启动防御代谢,同时激活COLD11介导的DNA修复,形成“感知-防御-修复”的完整闭环,让粳稻既能抵御低温损伤,又能修复已造成的损伤,最大化提升耐寒能力。
总结 粳稻耐寒的“全链条密码”,为育种提供新方向
种康院士团队的一系列关键研究层层递进,逐步解析了粳稻耐寒的核心分子机制,而这只是水稻耐寒复杂调控网络的一部分:
上游感知:粳稻特有的COLD1蛋白(SNP2ᴬ等位型)作为低温感受器,与RGA1互作,触发Ca²⁺信号,启动耐寒应答;
中游防御:COLD1调控叶绿体中维生素E/K1代谢及其他次级代谢,稳定细胞结构、清除活性氧,抵御低温损伤;
下游修复:COLD11通过GCG重复序列调控二聚体形成,高效修复低温诱导的DNA双链断裂,弥补损伤。
双绿源
上述分子机制的解析,为水稻耐寒育种提供了精准的分子靶点,也为分子标记辅助育种技术的应用奠定了基础。
双绿源依托这些核心研究成果,针对COLD1和COLD11等关键基因,开发分子检测标记,实现水稻耐寒性状的精准筛选与定向改良,为全球水稻抗逆育种提供高效技术支撑。





“绿色生态农业论坛”暨国家现代农业与···

芯片育种公司双绿源获隆平生物数千万元···

祝贺!联盟成员周俭民研究员获2023···
华中农大张启发院士/南京农大万建民院···

祝贺!联盟成员董莎萌教授

联盟秘书长张兴平博士在第七届国际瓜类···